
Au visionnage d’un documentaire animalier ou tout simplement au détour d’une promenade en forêt, vous avez sûrement déjà rencontré un animal aux caractéristiques extravagantes. Tellement extravagantes que c’est à se demander si cela ne va pas à l’encontre de la sélection naturelle. Prenez par exemple l’encombrante queue du paon, les immenses voiles des combattants ou bien les impressionnants bois des cerfs. Ce sont de belles ornementations, effectivement, mais aussi très embarrassantes pour se déplacer et très visibles par les prédateurs ! Une question se pose alors : comment de telles caractéristiques peuvent-elles être sélectionnées alors qu'elles semblent impacter négativement la survie des individus qui les portent ?
La réponse se cache dans un concept que je m’apprête à vous présenter : la sélection sexuelle. Dans la suite de cet article, nous nous demanderons pour quelles raisons dans la grande majorité des espèces, ce sont les femelles qui choisissent les mâles et comment elles les choisissent.
Puis, nous entrerons dans l’intimité d’autres animaux non-humains pour comprendre quelques stratégies sont sélectionnées au cours de l’évolution pour persuader les femelles de s’accoupler. Nous verrons ainsi que dans le monde animal il existe des tricheurs et que parfois la force et la beauté ne font pas le poids face à certains petits malins !
Ce qu’on retient du concept de sélection naturelle élaboré par Darwin et Wallace (plus de détails dans notre article à ce sujet), est que les individus possédant les caractéristiques les plus adaptatives, survivent mieux, se reproduisent mieux et donc sont sélectionnés de générations en générations. Seulement, si vous êtes un.e observateur.ice de la nature, vous devriez détecter une incohérence dans toutes ces histoires… En effet, lorsque l’on regarde attentivement le monde animal on s’aperçoit que les mâles et les femelles ont parfois des phénotypes différents. Les mâles sont souvent plus colorés ou dotés d'ornements voyants. Les femelles possèdent, en général, des attributs plus discrets. Au regard de la théorie de la sélection naturelle, il semble contre-intuitif que des individus soient sélectionnés alors qu’ils arborent des pelages multicolores qui les rendent plus visibles pour les prédateurs et les encombrent fortement.
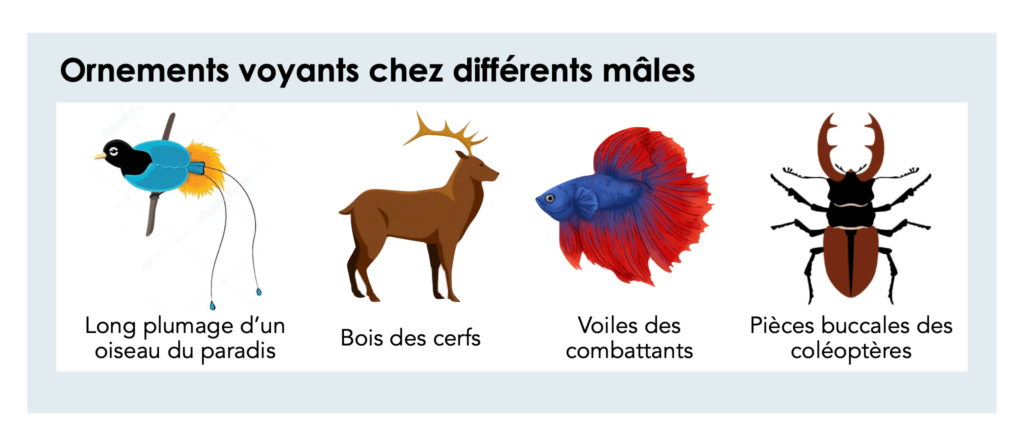
Mais alors, Darwin et Wallace se seraient-ils trompés ? À quoi bon favoriser des caractéristiques si handicapantes qui augmentent la probabilité de mourir ? Et bien la réponse réside tout simplement en une phrase : pour avoir accès à la reproduction !
Tous deux savaient que l'on ne pouvait s'attendre à ce que la sélection naturelle favorise l'évolution de traits désavantageux. Bien qu’ils eussent des idées divergentes à ce sujet ils ont quand même pu offrir une solution à ce problème : la sélection sexuelle. Ainsi dans son livre « The descent of man and selection in relation to sex »[1] paru en 1874, Darwin propose un nouveau processus évolutif différent de la sélection naturelle. Ce processus ne dépend pas d’une lutte pour l’existence par rapport à d’autres organismes ou à des conditions environnementales, mais d’une lutte entre les individus d’un sexe, pour la possession de l’autre sexe.
La sélection sexuelle est une théorie selon laquelle l'évolution de certains traits physiques remarquables tels qu'une coloration prononcée, une taille accrue ou des ornements frappants chez les animaux peut permettre à ceux qui les possèdent d'obtenir plus facilement des partenaires.
Mais pourquoi, dans la majorité des cas, ce sont les mâles qui se retrouvent parés d’ornements supplémentaires ?!
La théorie la plus répandue pour expliquer ce phénomène est l’anisogamie. « Anisos » signifie non-égal et « gamos » veut dire mariage. Ainsi, l’anisogamie désigne une forme de fécondation dans laquelle les gamètes des deux sexes diffèrent, notamment par la taille.
Les gamètes mâles (spermatozoïdes) sont beaucoup plus petits que les gamètes femelles (ovocytes). Chez les humains, un ovocyte mesure environ 120µm alors qu’un spermatozoïde mesure environ 50µm. De plus, il existe une différence de coût de production importante entre la production d’un gamète mâle et celle d’un gamète femelle. Les ovocytes vivent plus longtemps et contiennent des ressources nutritives, ils sont plus « chers » (métaboliquement) à fabriquer.
L’idée principale de cette théorie est que les mâles produisent des gamètes plus nombreux et peu coûteux. Les femelles n’en possèdent qu’un stock limité et précieux. De ce déséquilibre va découler un conflit sexuel. Les femelles ont intérêt à choisir avec attention un mâle alors que les mâles ont tout intérêt à féconder le plus de femelles possibles.
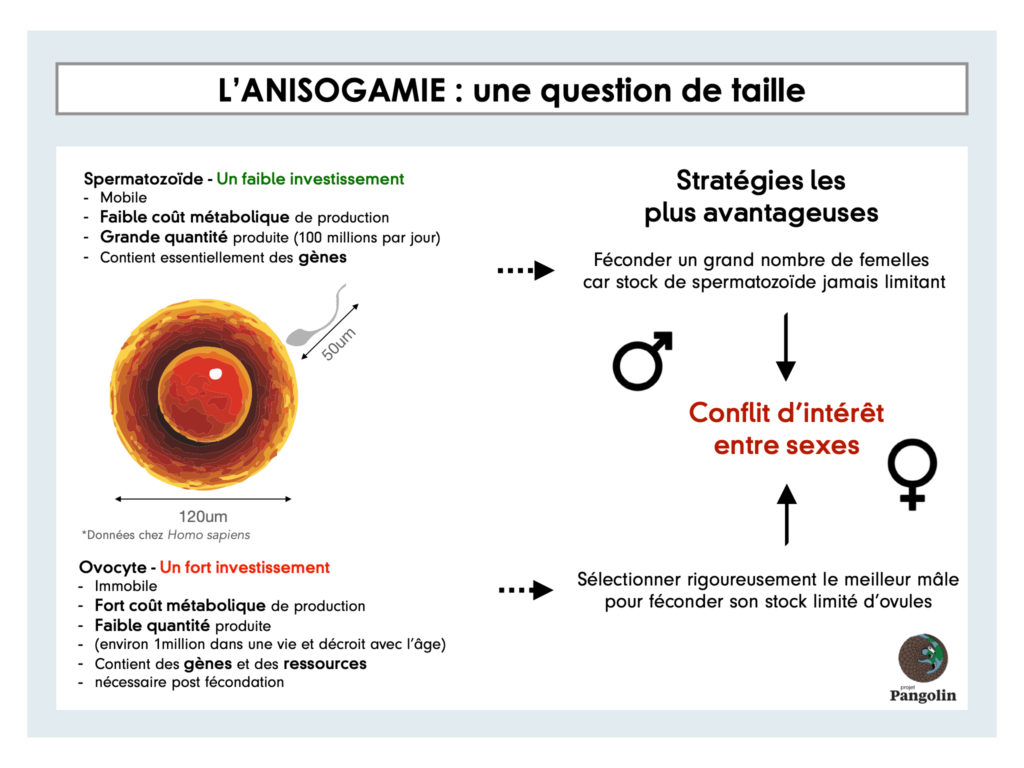
Au fil de l’évolution, cette guerre des sexes a mis les mâles en compétition entre eux pour accéder à une quantité de gamètes femelles limitée et ainsi s’accaparer la paternité tant espérée.
NB : il est intéressant de se demander comment l’anisogamie a émergé. Mais aussi pourquoi il existe des sexes différenciés. L’une des réponses réside dans le fait que cela permet d’éviter l’autofécondation. Si cela vous intéresse je vous invite à lire cet article en français qui porte sur le sujet.
Si les objectifs des mâles sont clairs. Ils ont besoin des femelles pour s'assurer une descendance et sont en compétition pour y accéder. Qu'est-ce que les femelles y gagnent et pourquoi choisir un critère plutôt qu’un autre ?
Dans certains cas c’est assez limpide. Si on prend pour exemple une femelle gorille qui aurait le choix entre un gorille gringalet et de faible rang social ou un gorille dominant et musclé, on s’imagine assez facilement pourquoi elle choisirait le gorille dominant. En effet, ce mâle a probablement accès à plus de nourriture et peut protéger son territoire. La femelle y gagne de la sécurité physique et alimentaire. Maintenant, si on prend l’exemple du paon, il est un peu moins intuitif de comprendre pourquoi les femelles préfèrent les mâles qui font le mieux la roue.
Le choix des femelles repose sur deux grandes catégories de critères : les avantages directs et indirects.
Un avantage direct va être immédiatement identifiable par la femelle et augmenter instantanément sa fécondité ou de sa durée de vie. Par exemple, les femelles peuvent obtenir : un meilleur accès à la nourriture, une protection contre les mâles qui les harcèlent ou une aide pour élever leur progéniture, ou encore éviter d'être infectées par des parasites (ou d'autres maladies) en choisissant des mâles en bonne santé.
Les avantages directs peuvent être de différentes natures. Certaines espèces offrent des cadeaux nuptiaux qui sont en général de la nourriture. On retrouve ce comportement chez une grande diversité d’organismes comme les oiseaux, les insectes et les araignées. Parfois il existe une corrélation positive entre la taille de l’offrande et le temps de copulation autorisé par la femelle. Ainsi si le mâle fournit une grosse proie à la femelle cela lui permettra de transférer plus de spermatozoïdes [3,4]. Chez d’autres espèces comme la Veuve noire à dos rouge, les mâles s’offrent eux-mêmes comme cadeau nuptial en se laissant dévorer [5].

Un autre avantage direct et qui paraît évident est de choisir un partenaire sexuel qui participera à l’élevage des jeunes pour certaines espèces. Les femelles choisiront un bon père pour les aider à construire un nid et à nourrir les petits. Ces comportements sont observés chez de nombreuses espèces d’oiseaux mais aussi chez les souris.
Un dernier exemple est retrouvé chez les grenouilles taureau. Les femelles vont préférer des mâles avec le « meilleur » territoire. Les œufs seront ainsi protégés des prédateurs pendant leur développement.
Un avantage indirect va, cette fois-ci, être moins évident pour la femelle et peut être interprété comme un pari. La femelle va devoir détecter si un mâle est un bon partenaire à partir d’indices plus ou moins clairs (pour nos yeux d’humains). Si sa mise est gagnante alors la femelle produira des descendants en bonne santé et capables à leur tour de produire de nombreux descendants, augmentant alors indirectement le succès reproducteur de la femelle.
Pour faire son choix, la femelle va se baser sur des signaux particuliers qui seront interprétés comme des indicateurs de qualité. Souvent ce sont des traits physiques exagérés qui ont été sélectionnés chez les mâles au fil de l’évolution. Ils sont appelés caractères sexuels secondaires. Deux théories complémentaires permettent d’expliquer l’apparition de ces drôles de caractéristiques.
La théorie de l’emballement de Fisher propose qu’au fil des générations, un trait non-adaptatif, visible et garant d’une bonne qualité va être sélectionné par les femelles. S’en suivra alors un mécanisme de boucle de rétroaction positive. Les femelles sont de plus en plus intéressées par cette caractéristique alors la sélection sexuelle va favoriser son augmentation.
Par exemple les femelles se verront choisir les mâles avec les plus longs plumages car leurs descendants hériteront aussi d’un long plumage et seront à leur tour choisit par les femelles de la génération suivante, maximisant alors leurs probabilités de dispersion de gènes (Fisher 1930).
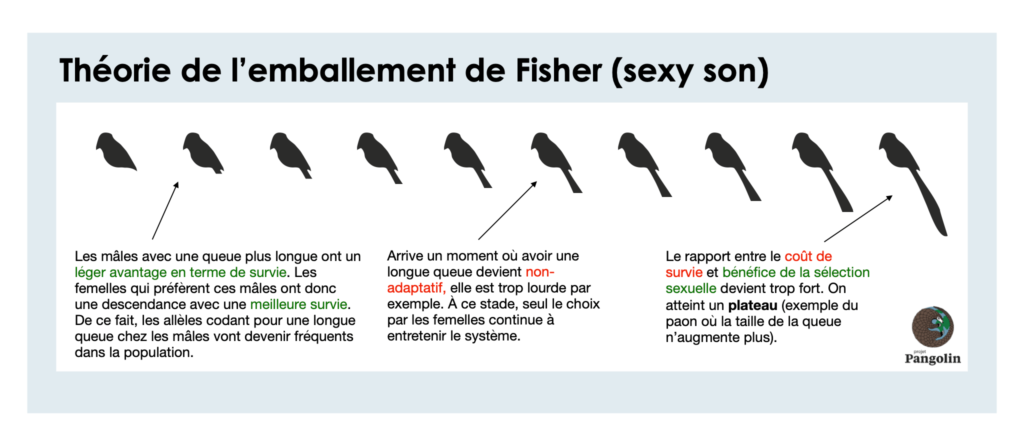
Ce mécanisme entraînera alors une sélection effrénée pour l'exagération de l'ornement et de la préférence. Ce mécanisme se poursuivre jusqu'à ce que les coûts de production de l'ornement dépassent l'avantage reproductif de sa possession. En théorie il peut s’agir de n’importe quel trait non-adaptatif tant que les femelles peuvent facilement les comparer. Le coût n’apparaît qu’au stade final d’un emballement évolutif.
La théorie alternative à l’emballement fisherien se nomme la théorie du handicap. Elle est aussi appelée la théorie des bons gènes.
Selon cette théorie du handicap, les mâles arborant des traits extravagants voire dangereux pour leur survie seraient des mâles de bonne qualité globale. En effet, les mâles handicapés par un trait extravagant ont été capables de survire dans un environnement hostile alors même qu’ils devaient (par exemple) trainer une immense queue multicolore (Zahavi 1975). Leur caractère extravagant serait alors un signal honnête informant sur leurs conditions physiques et donc leurs ‘bons gènes’.

Tous ces mécanismes reposent sur l’existence supposée de gènes conférant une meilleure vigueur (hypothèse des « bons gènes »), révélés par des indices extérieurs handicapants et ne pouvant donc être développés que par les individus de très bonne qualité individuelle. Cependant, les preuves du choix des femelles pour les bons gènes restent rares malgré des décennies d'études sur le choix du partenaire chez de nombreux organismes. Ce manque apparent de consensus continue de créer un débat quant à l'importance du modèle des bons gènes dans le domaine de la biologie évolutive.
La plupart du temps lorsque l’on parle de sélection sexuelle on pense aux affrontements à l’approche de la saison des amours ou bien aux somptueuses parades nuptiales. Dans le règne animal ces stratégies sont très répandues et surviennent avant la copulation. Si ces comportements ont été sélectionné au cours de l’évolution c’est principalement car ils permettent de départager les meilleurs mâles. À la fois en termes d’avantage indirect (si je choisis le mâle qui gagne, alors ma progéniture héritera probablement elle aussi des gènes qui lui permettront de gagner la compétition à son tour), ou bien d’avantage direct (si je choisis le mâle le plus fort, alors j’augmente ma probabilité de survie et celle de ma progéniture car je serais mieux protégée des prédateurs).

Les femelles ont parfois le choix de ne pas mettre tous leurs œufs dans le même panier. Chez ces espèces il est alors fréquent qu’elles s’accouplent avec différents mâles pour maximiser leurs chances d’avoir une descendance en bonne santé. Dans cette situation, la compétition entre mâle intervient au niveau du sperme et s’appelle la compétition spermatique.
Les mâles n’ont pas intérêt à ce que d’autres mâles viennent féconder la femelle avec laquelle ils viennent de s’accoupler. Ainsi, au cours de l’évolution de multiples stratégies ont vu le jour à différentes temporalités du stade copulatoire.
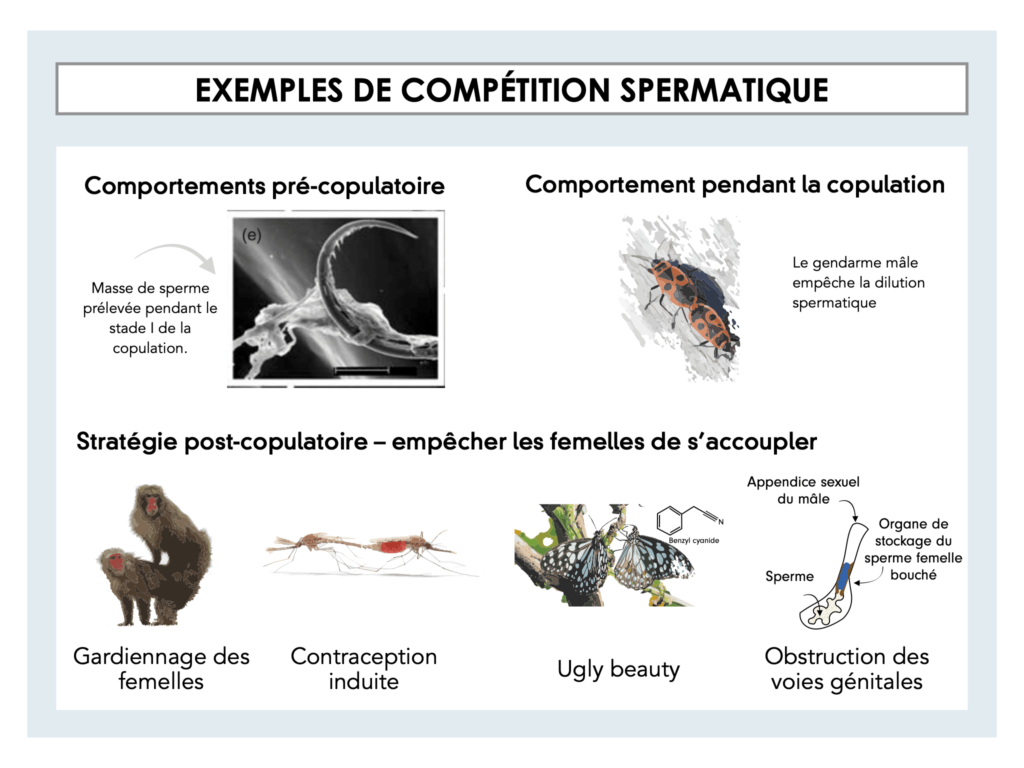
Chez certaines espèces (comme les libellules par exemple) lorsque la femelle a déjà été fécondée, il arrive que le mâle suivant prenne plusieurs heures pour nettoyer la cavité spermatique de la femelle avant d’y introduire sa semence. Ainsi, à l’aide de petits plumeaux, il élimine le sperme d’éventuels prédécesseurs et accroît ses propres chances de paternité.
Afin de prolonger la durée de copulation, certains mâles ne vont plus lâcher la femelle d’un pouce. L'objectif étant de maximiser les chances de fécondation de leurs spermatozoïdes pendant l’accouplement. Pour cela il arrive que certains mâles fassent office de bouchon copulatoire. Chez certains phasmes le contact génital peut durer jusqu’à 79 jours ! [10].
Obstruction des voies génitales : Certaines espèces (blattes, criquets, et autres insectes) vont accompagner leur sperme d’une substance (glandes à ciment) destinée à obstruer les voies génitales de la femelle lors d’accouplements ultérieurs (par exemple, la mouche du fumier).
Contraception induite : Une autre stratégie consiste à libérer une substance qui empêche tout accouplement ultérieur. Chez les moustiques, les mâles produisent une substance par le biais de leur glande sexuelle secondaire qui inhibe la réceptivité des femelles inséminées. Ainsi, elles refusent la plupart des accouplements avec d'autres mâles [11].
Gardiennage des femelles : D’autres vont simplement ne plus quitter les femelles des yeux et éloigner tous mâles susceptibles de vouloir féconder la femelle [3], c’est le cas de certains singes ou bien des canards colvert.
Ugly beauty : Une autre tactique consiste à rendre les femelles peu attirantes pour les autres mâles. Ce phénomène est particulièrement bien connu chez les papillons de nuit. En effet, les mâles déposent des anti-aphrodisiaques sur les femelles lors de l’accouplement [3].
Jusqu’à présent nous avons vu que les caractères sexuels secondaires (par exemple, la queue du paon) étaient des indicateurs honnêtes de la qualité sous-jacente des mâles. Les coûts associés à la production et à l’entretien de ces caractéristiques extravagantes informaient les femelles sur l’état de santé global des mâles qui les portaient. En effet, plus un mâle est en bonne forme physique, plus il a accès à des ressources énergétiques de qualité. Et plus il est capable de les gaspiller en se pavanant avec une très longue queue dans la forêt. Certains mâles y ont vu l’opportunité d'optimiser leur allocation d’énergie en réduisant certains coûts liés aux traits sexuels secondaires sans nuire à leur succès reproductif global. C’est là que la triche rentre en jeu !
Certains mâles araignées remplissent leurs cadeaux nuptiaux de soie à la place d’une proie pour duper les femelles et se reproduire plus longtemps [14].
A défaut de ne pouvoir rivaliser avec les mâles plus imposants, certains vont ruser pour accéder tout de même à la fécondation. Chez certains batraciens, les mâles plus petits vont suivre de près les gros mâles et agripper les femelles attirées à ce dernier.
Les mâles d’autres espèces de poissons vont profiter de la fécondation qui se fait par jetée de sperme dans l’eau pour mêler le leur à celui d’autres poissons. Chez certains scarabées, les mâles vont aller féconder les femelles en cachette tandis qu'un mâle plus imposant va monter la garde.
Enfin, de nombreuses femelles choisissent des mâles qui vont accorder tout leur temps pour aider au nourrissage ou bien défendre le territoire. Les mâles se retrouvent ainsi tiraillés entre rester avec la femelle et féconder le plus de femelles possibles. Ainsi, il n’est pas rare d’observer des mâles aller féconder d’autres femelles en douce. Une étude sur des oiseaux, les Parulines orangées a ainsi montré que près de 27% des progénitures ont été identifiées comme résultant d'un accouplement extra-pair [15] !

Historiquement le domaine de la sélection sexuelle reste très jeune. En effet, la recherche dans ce domaine n’a démarré que dans les années 1960 [22]. Il n’est donc pas étonnant que le concept de sélection sexuelle reste méconnu, alors que le grand public a déjà connaissance de la sélection naturelle.
La science n’est pas neutre. Un.e scientifique est pris.e dans une matrice axiologique dont iel ne peut se défaire et dont il n’est pas souhaitable qu’iel se défasse car cela enlèverait à la science sa dimension humaine. Il est faux de penser que lorsqu’un.e scientifique pose son chapeau, son manteau et enfile sa blouse blanche iel efface le filtre interprétatif entre nous et le réel.
Il faut donc rester prudent quant aux observations que nous faisons sur le monde qui nous entoure, bien que nous puissions avoir des études très sérieuses sur la question, il n’en reste pas moins que notre perception des choses est subjective, spécialement dans un monde aussi complexe que le nôtre. L’idée de valeur commune est difficile à maintenir. Nous avons donc, aujourd'hui plus que jamais, besoin de partager les connaissances et les points de vue d'une diversité de chercheur.e.s sur un sujet de recherche car c’est en ce sens que nous pourrons diminuer au maximum nos interprétations personnelles.
Nous venons dans cet article de voir quelques exemples que j’ai choisis, mais en réalité il existe autant de différences qu’il n’existe d’espèces. Je ne vous ai pas parlé des espèces sans reproduction sexuée ou bien encore celles chez qui le sexe change au cours de la vie des individus (ce n’est que partie remise) ! Je reste personnellement toujours fasciné par toutes ces singularités qui nous entourent d’autant que la sexualité en générale façonne et occupe une place primordiale dans les processus évolutifs : celui de la passation des gènes !
1- Darwin, C., 1875. The Descent of Man, and Selection in relation to Sex. Nature, 11(277)
3- Albo, M. and Peretti, A., 2015. Worthless and Nutritive Nuptial Gifts: Mating Duration, Sperm Stored and Potential Female Decisions in Spiders. PLOS ONE, 10(6)
4- Gwynne, D., 2008. Sexual Conflict over Nuptial Gifts in Insects. Annual Review of Entomology, 53(1)
5- Andrade, M., 1998. Female hunger can explain variation in cannibalistic behavior despite male sacrifice in redback spiders. Behavioral Ecology, 9(1)
6- Toft, S. and Albo, M., 2016. The shield effect: nuptial gifts protect males against pre-copulatory sexual cannibalism. Biology Letters, 12(5)
7- Cecie Starr and Ralph Taggart, Biology – the Unity and Diversity of Life, 6th Ed., Wadsworth Publishing Company, 1992
8- Edward T. Hall, The Hidden Dimension, New York, Anchor Books, 1966
9- Córdoba-Aguilar, A., Uhía, E. and Rivera, A., 2003. Sperm competition in Odonata (Insecta): the evolution of female sperm storage and rivals' sperm displacement. Journal of Zoology, 261(4)
10- Gangrade, G., 1964. Accessory glands of female phasmid, Necroscia sparaxes Westwood. Annals and Magazine of Natural History, 7(84)
11- Helinski, M., et al., 2012. Duration and dose-dependency of female sexual receptivity responses to seminal fluid proteins in Aedes albopictus and Ae. aegypti mosquitoes. Journal of Insect Physiology, 58(10)
12- Parker, G., 1970. Sperm competition and its evolutionary consequences in the insects. Biological Reviews, 45(4)
13- van Noordwijk, A. and de Jong, G., 1986. Acquisition and Allocation of Resources: Their Influence on Variation in Life History Tactics. The American Naturalist, 128(1)
14 - Ghislandi, P., et al., 2017. Silk wrapping of nuptial gifts aids cheating behaviour in male spiders. Behavioral Ecology, 28(3)
15 – Morton Catherine Heidrich Virginia., 2013. PhD thesis. Who's Your Daddy? A Study of Extra-Pair Copulation and Mating Behaviors of Protonotaria citrea Behaviors of Protonotaria citrea.
16 - Sinervo, B. and Lively, C., 1996. The rock–paper–scissors game and the evolution of alternative male strategies. Nature, 380(6571)
17- Puts, D., 2021. Human sexual selection. 20015. Current Opinion in Psychology.
18- Scott, I., et al., 2014. Human preferences for sexually dimorphic faces may be evolutionarily novel. PNAS, 111(40)
19- Wheatley, J.,et al., 2014. Women’s faces and voices are cues to reproductive potential in industrial and forager societies. Evolution and Human Behavior, 35(4)
20- Collins, S. and Missing, C., 2003. Vocal and visual attractiveness are related in women. Animal Behaviour, 65(5)
21- Singh, D., et al., 2010. Cross-cultural consensus for waist–hip ratio and women's attractiveness. Evolution and Human Behavior, 31(3)
22- Giraldeau, L., Cézilly, F. and Danchin, E., 2005. Écologie comportementale. Paris: Dunod.
